Mechanistic distribution models
One research focus is developing dynamic bioenergetic models that integrate individual energetics and population dynamics to predict species' distributions. The models are among the first to produce dynamic range predictions based on first principles of morphology and physiology. Research, in collaboration with Joan
Roughgarden at Stanford, developed and field tested the models for Caribbean lizards along elevation gradients (Buckley and Roughgarden JAE 2005, Buckley and Roughgarden FEC 2006).
Research at NCEAS and the Santa Fe Institute has extended the dynamic bioenergetic modeling framework to predict current ranges for North American lizards and those following climate change. The analysis highlights the
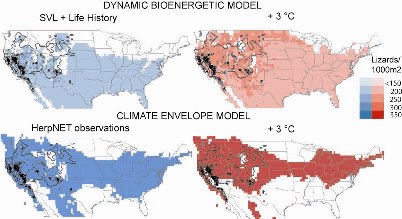 limitations of correlative approaches to species' ranges and the importance of considering physiology and population
dynamics. Recent research (Buckley AmNat 2008) investigates the range implications of geographic variation in traits. The dynamic model predicts individualistic responses to a uniform 3 ° C warming, but a northward shift in the northern range boundary for all populations and species. (Figure: Predictions of the dynamic bioenergetic model and a climate envelope model for the western fence lizard, Sceloporus graciosus, Buckley AmNat 2008)
limitations of correlative approaches to species' ranges and the importance of considering physiology and population
dynamics. Recent research (Buckley AmNat 2008) investigates the range implications of geographic variation in traits. The dynamic model predicts individualistic responses to a uniform 3 ° C warming, but a northward shift in the northern range boundary for all populations and species. (Figure: Predictions of the dynamic bioenergetic model and a climate envelope model for the western fence lizard, Sceloporus graciosus, Buckley AmNat 2008)
Mechanistic range models enable incorporating biotic constraints on species ranges such as trait evolution, species interactions, and dispersal limitations. We participate in a

 joint NCEAS and NESCent working group (PIs: M. Angilletta, L. Buckley, R. Holt, and J. Tewsbury) that is bringing together ecologists, evolutionary biologists, phylogeographers, and physiologists to extend mechanistic range models to include these biotic constraints. The working group members apply theoretical, lab, and field based approaches and work on taxa including plants, insects, reptiles, amphibians, birds, mammals, and intertidal marine organisms. Extending mechanistic range models to other taxa, including endotherms, will enable comparing the energetic constraints on ectotherms and endotherms.
joint NCEAS and NESCent working group (PIs: M. Angilletta, L. Buckley, R. Holt, and J. Tewsbury) that is bringing together ecologists, evolutionary biologists, phylogeographers, and physiologists to extend mechanistic range models to include these biotic constraints. The working group members apply theoretical, lab, and field based approaches and work on taxa including plants, insects, reptiles, amphibians, birds, mammals, and intertidal marine organisms. Extending mechanistic range models to other taxa, including endotherms, will enable comparing the energetic constraints on ectotherms and endotherms.
Energetic and ecological constrains on reptile abundance
Other recent research uses global scale datasets to examine energetic and ecological constraints on abundance, diversity, and species turnover. We found that lizard population densities decrease with energetic needs and increase with energy supply
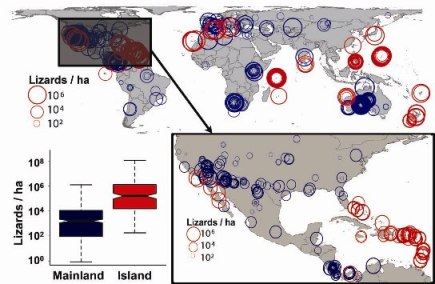 (Buckley and Jetz Ecology 2008). While energetic constraints act consistently across mainlands and islands, local abundances are one order of magnitude higher on islands than on mainlands due to release from predation and competition (Buckley and Jetz ELE 2007). Our results demonstrate the dramatic effect insularity has on the interplay between biotic and abiotic control of animal abundances. This highlights the elevated sensitivity of island biota to small ecological perturbations such as invasions or losses of even single species.
(Figure: The order of magnitude higher lizard density (lizards/ha, with larger circles indicating higher density) on islands (red) compared to mainlands (blue), Buckley and Jetz ELE 2007.)
(Buckley and Jetz Ecology 2008). While energetic constraints act consistently across mainlands and islands, local abundances are one order of magnitude higher on islands than on mainlands due to release from predation and competition (Buckley and Jetz ELE 2007). Our results demonstrate the dramatic effect insularity has on the interplay between biotic and abiotic control of animal abundances. This highlights the elevated sensitivity of island biota to small ecological perturbations such as invasions or losses of even single species.
(Figure: The order of magnitude higher lizard density (lizards/ha, with larger circles indicating higher density) on islands (red) compared to mainlands (blue), Buckley and Jetz ELE 2007.)
Despite ongoing population declines and potential future susceptibility to environmental change, the broad-scale ecology of reptiles and amphibians has received relatively little attention. Recent research examined environmental and historical constraints on global patterns of amphibian richness (Buckley and Jetz PRSb 2007). The analysis is the first for a vertebrate ectotherm clade at the broad scale. Ongoing research is examining other broad scale patterns of vertebrate ectotherm distribution. Comparing the broad scale ecologies of ectotherms and endotherms addresses how the groups' differential use and acquisition of energy influences patterns of abundance, distribution, and diversity.
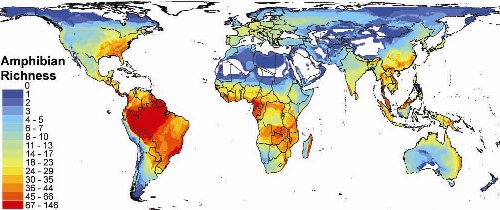
(Figure: The global pattern of amphibian richness, Buckley and Jetz PRSb 2007.)


