March 26-28, 2007
Embryology Biology 441 Spring 2007 Albert Harris
Embryology of the Nervous System
An individual nerve cell is called a neuron .
Other cells of the central and peripheral cells are called glia, or glial cells.
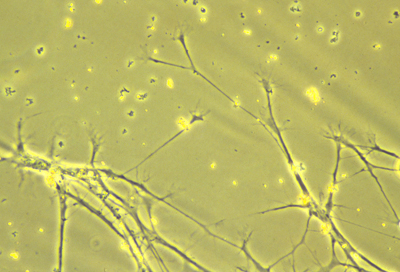
nerve axons
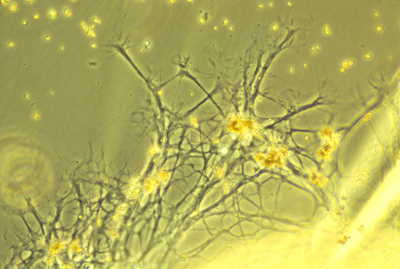
axons and glia
One example of a kind of glial cell are Schwann cells. These wrap parts of their plasma membrane around nerve axons, to form the myelin sheath (in the peripheral nervous system).
Schwann cells are derived from neural crest cells.
Inside the brain and spinal cord, there are several other kinds of glial cells, such as "oligodendrocytes".
Nerve fibers are long strands of cytoplasm and plasma membrane. The longer nerve fibers are called axons; and most short nerve fibers are called dendrites.
I used to assume that the difference between axons and dendrites must only be size. But it has been discovered that the microtubules inside axons are all oriented with their plus ends in the same direction, away from the cell body; whereas inside dendrites some of the microtubules are oriented with the plus ends away from the cell body, and others oriented in the opposite direction.
Both axons and dendrites are formed by active "amoeboid" crawling locomotion of special areas of cytoplasm at their tips called "growth cones". Axon elongation is a special case of crawling locomotion, not simply growth. Tissue culture was invented by Ross Harrison to prove this fact.
"Motor nerves" carry nerve impulses from some part of the central nervous system to muscle cells (or sometimes to gland cells), and stimulate the muscles to contract. The motor nerve cell bodies are located inside your spinal cord.
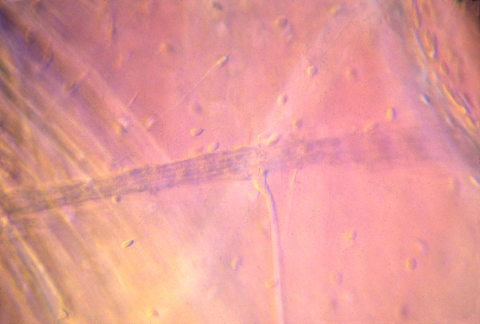
nerve innervating muscle in tadpole
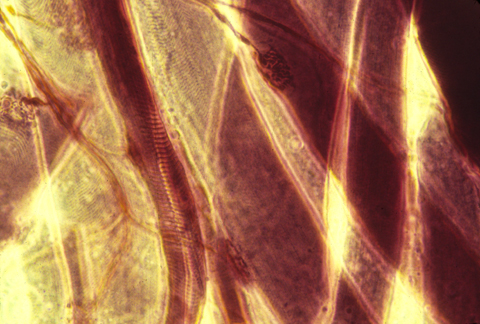
nerve-motor synapses
Autonomic nerves are a special kind of motor nerves, that control involuntary processes like the rate the heart beats, and intestinal functions
"Sensory nerves" carry nerve impulses from some part of the periphery, back to the spinal cord, or sometimes directly to the brain. Sensory nerve cell bodies are located in a series of spinal sensory ganglia, one pair per somite, next to your spinal cord.
Notice how long the axons have to be that stimulate your foot and hand muscles, and that sense touch, pain etc. in your hands and feet.
These axons have to reach all the way from between your shoulders out to your hands, and all the way from your lower back, down to your feet.
So we are talking about cells that are 2 or 3 feet long, but only a micron, or up to 5 or 10 microns in diameter.
{In case you are interested, those giant axons in squids are multi-nucleate, and formed by fusion of thousands of original cells.}
The neurons in our bodies are all unicellular. Nerve cells never undergo mitosis, or even synthesize DNA, after they have started making an axon.
If some nerve axons are cut in your arm, or leg or back, then the cut end will re-form a "growth cone", which will resume active locomotion, and sometimes find its way all the way back to the original muscle cell, and re-form a synapse or crawl back to the original location in the skin.
But in the embryo, they hadn't needed to crawl nearly as far, to reach their correct connections.
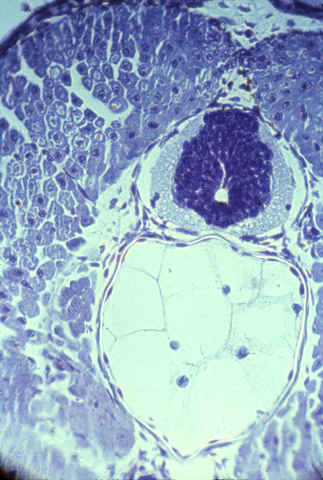
The brain and spinal cord are made of nerve cells, axons, and glial cells tightly packed together. The following photograph shows a histological cross section of the spinal cord of a salamander. The "white matter" is lots of nerve axons packed together. The blue-stained nuclei surrounded by the white matter shows the location of a mass of nerve cell bodies (this is called the "grey matter" because of its grey color when not dyed.
The human central nervous system is also subdivided into grey matter and white matter, except that in brains of higher animals, the grey matter is on the outer surface, and the white matter more interior. The following are histological sections of parts of a mammal brain, stained (dyed) with silver metal in a way that colors only a random 1 or 2 % of the cells, but colors all of the cell body, axons and dendrites of this minority of cells that are stained. Without this method, the cellular structure of the central nervous system would never have been mapped. That is because the nerve cells and fibers are packed together so tightly that staining them all would produce a solid mass of color.
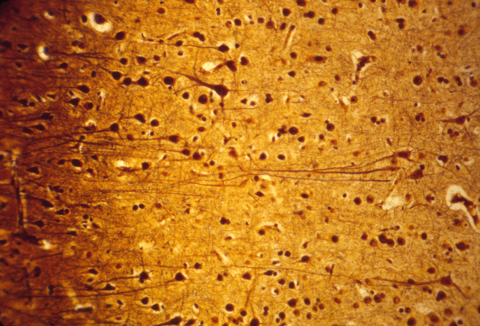
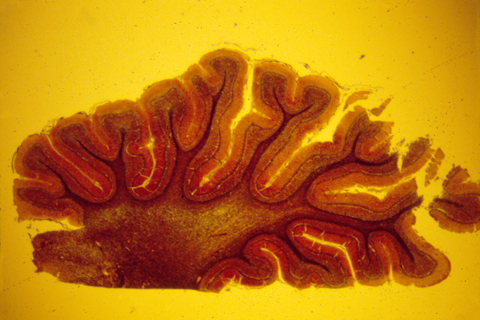
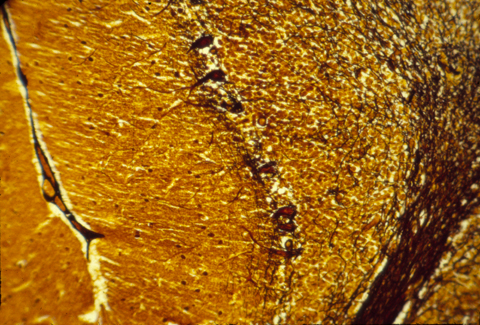
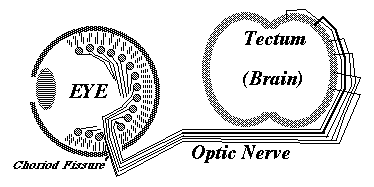
Much research has been concentrated in the mechanisms that guide the growth cones of the developing optic nerve from the neural retina of the eye, back to the particular part of the brain that it innervates. The retinal cells whose axons form the optic nerve are called "ganglion cells", you have about a million of them in each eye, and each one sends of single axon back to the opposite side of the brain (mostly!).
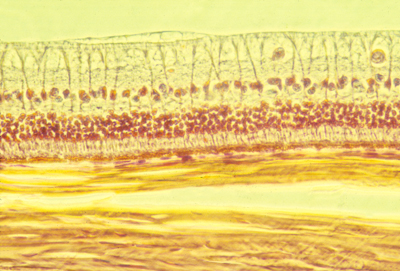
Histological section of the retina of a rabbit. The light from the pupil and lens comes from the top (as this section is oriented), passes through the optic nerve axons at the top, and the large ganglion cell nuclei below and among them, and on downward to the rod and cone cells in the middle. Below then is the pigmented retina, and below that you can see thick strong sheets of collagen fibers that are wrapped around the eyeball and give it strength.
In birds, amphibians and fish, the part of the brain they innervate is called the "optic tectum". At the tectum, the growth cones of the optic nerve axons spread out over the brain surface is the same geometric pattern as the parts of the retina where they are coming from.
They then form synapses in what amounts to an up-side down and backwards (reversed back to front) "map" of the parts of the retina where their ganglion cells are located. This pattern of connection is called a neural projection.
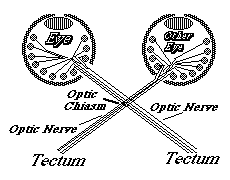
The retino-tectal projection is one among dozens, or probably hundreds of different neural projections in our nervous system. For example, the sensory nerves of the whole surface of the body form a neural projection to a strip along the roof of the forebrain; And the motor nerves for all the voluntary muscles of the body form a neural projection from another strip of the roof of the forebrain, side by side with the sensory projection, separated by a certain infolding of the roof of the brain.
The causal mechanism for the formation of the retinotectal connection has long been thought to be some kind of adhesive specificity. The first demonstration of this was about 35 years ago.
The current belief is that special proteins called ephrins occur in tectal cells, where they vary in number per cell according to a gradient (notice this is not a diffusion gradient). Another kind of cell surface receptor, called an ephrin receptor, is present on the surfaces of the ganglion cells and their axons. Ephrin proteins bind selectively to ephrin receptor proteins; and the higher the concentration of ephrins, then the lower the concentration of ephrin receptors needed to produce an the essential effect - but this effect (surprise!) is not adhesion between axon and brain cell, but somewhat the reverse!
Ephrin binding inhibits further locomotion of a growth cone, often resulting in its retraction a short distance. The basic idea is that this is super-imposed on some less specific attraction of optic nerve growth cones, with inhibition of crawling being produced by ephrin-ephrin receptor binding. When the farthest part of the tectum has he higher concentration of ephrin, then only the axons with the lowest concentrations of ephrin receptors will be able to get that far before further crawling is blocked. Axons with the highest concentrations of ephrin receptors will be stopped at the lower side of the tectum, where the brain cells have the highest concentrations of ephrin.
There are two classes of ephrins and receptors, ephrin A and ephrin B, and the evidence is that one controls the dorso-ventral axis and the other controls the anterior-posterior axis. There is still much to be learned however.
Some other kinds of evidence suggests that ganglion cell axons interact with each other. For example, Martha Constantine Paton did experiments in which an extra eye was grafted onto the side a frog head, and the pattern of innervation by their ganglion cells was studied in the tectum. Instead of the nerves from both eyes connecting in a homogeneous mixture, they formed stripes. Each stripe of the tectal surface became innervated mostly by axons from one or other of the eyes, and the adjacent stripes on either side of that stripe became innervated mostly by axons from the other eye.

GREETINGS, EARTH PEOPLE