January 14th lecture
Embryology Biology 441 Spring 2008 Albert Harris
Genes →→→→→→→→ cause →→→→→→ Anatomy
Cell differentiationplus Cell movements (active "amoeboid" rearrangements)
Mechanisms that cause and control cell differentiation:
-
* "Turn on" one subset set of luxury genes; Using transcription factors.
* "Turn off" all other subsets of luxury genes.
åCell differentiation is usually very stable (some mechanisms of self-perpetuation?)
-
* Plant cell differentiation tends to be less irreversible.
* If you could make human cells de-differentiate, and then re-differentiate into other cell types, that would be better than embryonic stem cells.
Tissue culture cells usually seem to dedifferentiate; but this is only apparent;
It's difficult to get them to re-differentiate, especially into different cell types.
(But there is a lot of research on how to do this.)
========================================================================
#1) Spatial control of cell differentiation, so they FORM AT the right places.
#2) Active rearrangements of differentiated cells, so they MOVE TO the right places.
(Both these methods are used; about half and half; but the first gets most attention.)
Sponges and Hydra use almost entirely the second of these methods.
Their differentiated cells constantly rearrange themselves.
This ability to rearrange by cell type was discovered 100 years ago at UNC, by H. V. Wilson, first in sponges and later in corals. ("Cell Sorting")
(Wilson concluded that the cells must be re-differentiating,
rather than rearranging, and spent 30 years trying to prove that.)
"Cell sorting" was later shown to occur by frog and salamander embryo cells, and then by bird and mammal embryo cells. This led to discovery of selective adhesion proteins on the outside surfaces of cells; examples include N-CAM and cadherins.
Probably there are many more causal mechanisms that cause and control cell sorting,
But selective adhesion proteins have satisfied almost everybody's curiosity,
Because it is a very "chemical" mechanism.
Later we will talk about Malcolm Steinberg's "Thermodynamic mechanism",
which the textbook likes and gives several pages to.
Other scientists (including Wayne Brodland and me) have published different theories, based on active cell contraction, instead of maximization of cell-cell adhesion.
Can you invent experiments that could discover which explanation is true?
These are questions that I hope to do experiments on during the next year.
*******************************************************************************
"Determination" means commitment to of a cell to differentiate into a certain cell type.
"Stem cells" used to mean cells that remained determined while dividing, and continually supplying certain kinds of differentiated cells (as bone marrow cells do).
The new meaning of stem cell is what we used to call un-determined cells.
These used to be called "totipotent cells"
"Determination of early embryonic cells occurs in a certain branching pathway, analogous to railway switches:
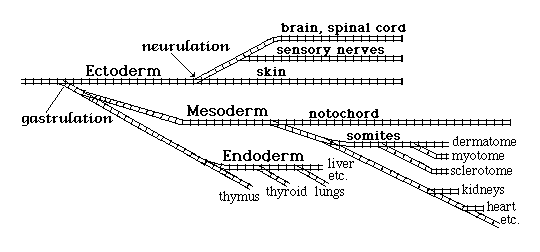
The first three branches are called the ectoderm, the mesoderm and the endoderm.
Not to be confused with Dur'm, which is a nearby city.
Ectoderm becomes nerves and the outer layer of the skin, etc.
Endoderm becomes digestive tract, lungs, and many glands.
Mesoderm becomes skeleton, muscle, kidneys, blood vessels, etc.
&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&
Concepts and theories about how differentiation is spatially controlled:
Reaction-Diffusion systems
Suppose there are two chemicals (which could be proteins, or ions, or anything)(Actually, they can be almost any quantitative variables)
Call one chemical "A" and the other chemical "B"
People tend to call one "The Activator" and the other "The Inhibitor",
but that just narrows your thinking.
-
1) Suppose A stimulates formation (or release) of both A and B.
2) Suppose B stimulates destruction (etc.) of both A and B.
(that's not very grateful of it!)
3) Suppose B diffuses faster than A.
(Or in any way acts at longer distances, sooner)
????????????????????????????????????????????????????????????????????????????????
This example illustrates the need for computer simulations, because almost nobody's intuition is strong enough to visualize the result.
But even I have written computer programs that show what will happen.
You don't really need to know all the variables and equations obeyed by a system in order to write a computer simulation.
And it's an unfair criticism to complain that variables in a program can be adjusted to make it produce whatever result you want to get!
The point it to use programs experimentally, to find out what variables and equations will be able to produce results that look like biological phenomena, And then use the programs to predict what other results you should test for.
!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
Alan Turing invented "reaction-diffusion" systems.
Scott Gilbert seems to think they only magnify pre-existing irregularities.
Notice his drawing of how he thinks they behave, in the textbook.
I don't think those drawings were really produced by any computer simulation (because I digitized his curves, and couldn't get any simulation to behave like them. Real simulations produce can produce better spacing.)