Embryology Biology 441 Spring 2009 Albert Harris
Lecture Notes and questions for Jan 12 and 14
Embryology is the study of the mechanisms that form the body, including all its anatomical structures and tissues.Two main processes build the body: cell differentiation and cell locomotion
1) Cell differentiation.
For example, some cells differentiate into skin cells,
other cells differentiate into nerve cells, & there are lots of different kinds of nerve cells.
The human body is made of about 250 kinds of differentiated cells.
Sponges have maybe ten, or fewer, differentiated cell types.
Dictyostelium, a "slime mold" has 4 cell types.
Eventually, researchers will be in a position to count and enumerate exactly
how many cell types each kind of animal has; but there may be gray areas.
(but there isn't much motivation to find out exactly how many)
* Differentiation consists of selective transcription of a different particular
sub-set of the approximately 30,000 genes.
(30 years ago, the estimated total number was 50,000 to 100,000.
The estimate has been decreasing, like the stock market.)
* Some genes are "expressed" (meaning that their protein gets synthesized)
in all or nearly all differentiated cell types: for example, genes for enzymes
used in basic metabolism, or for microtubule proteins, etc.)
These are called "housekeeping genes" (and housekeeping proteins)
Genes that are only expressed in certain differentiated cell types are called
"luxury genes" ( & luxury proteins). This wasn't such a good word choice,
but we are stuck with it, and I don't have any better suggestion.
But don't conclude that animals could survive without their luxury proteins.
* The main mechanism of differentiation is selective transcription, controlled by selective binding of special proteins ("transcription factors") that fit selectively to special DNA sequences ("promoter regions") that are a few base pairs upstream of the DNA that codes for the protein ("the structural gene")
Later in the course, you will learn a lot about some particular transcription proteins
and how they control body patterns in flies, which is a very hot area of research.
When researchers studied fly mutations that blocked embryonic development past
Different particular stages of development, most of the genes they discovered turned
out to code for transcription factors. Mutations in certain transcription factor genes
can cause organs to develop in the wrong place, where a different organ should form.
Those are called homeotic mutations. An important kind of genes that can do this are
the "hox genes".
Please understand that if DNA broke and rejoined at the wrong place, between the promoter region of one luxury gene and the coding sequence (structural gene) of some different gene, then this other gene would be expressed (get transcribed, and its protein get made) when the cell tried to make whatever protein is normally just downstream of that promoter region.
An example is that many subtypes of the kind of cancer called non-Hodgkin's Lymphoma are caused by accidental breakage and rejoining of DNA just upstream of genes that code for antibody proteins.
Questions for class discussion:
a) Why would this particular translocation only cause cancer of lymphocytes?(The exact same mutation in any other cell type would have no effect)
b) The genes moved downstream of the antibody gene promoter region
would need to code for proteins whose effects were of what kinds?
(That is, in order for cancer to result from the gene translocation?)
c) If you insert a gene that produces a visible or special-colored protein just downstream of a certain promoter region, then what will happen?
d) Do you know what's meant by a "reporter gene"?
e) Do you know what GFP is?
2) Cell locomotion ("cell motility", "Morphogenetic cell movements", and also a few other kinds of mechanical force exertion, which include active bending of epithelial cell sheets, inflation of fluid-filled cavities, and osmotic swelling of cartilage).
Growth has turned out to be much less important than everyone expected
as a cause of shape formation in animal development.
(Growth either in the sense of cell divisions or cell enlargements)
[NOTE: Higher plants don't have cell locomotion, they have osmotic swelling and cell divisions, but cells just differentiate at a certain place and stay there.]
Animal embryos have several sets of active cell rearrangements:
The first set of cell rearrangements is called gastrulation. during which the future digestive tract cells move from surface to interior and future muscle, bone, kidney etc. cells move from surface to interior.
Just about all kinds of animals have their own version of gastrulation.
Vertebrate embryos have a second major cell rearrangement,
called neurulation, which moves future nerve cells into the interior.
(but don't expect to see a sea urchin embryo neurulate)
How odd that future nerve cells start out on the outside surface, and then fold or crawl into the interior, before differentiating as nerves.
QUESTION: f) What do you think it means about control mechanisms that if you prevent them from moving inside (if you prevent neurulation) then they will differentiate into skin cells, instead of nerve cells
Everyone starts out as one cell: the fertilized egg cell.
This cell then divides into two, etc.
In mammal embryos, these divisions are NOT synchronous;
But in embryos of frogs, fish, and many others, divisions are synchronous:
1 cell, 2 cells, 4 cells, 8 cells, 16, 32, etc.
Synchrony has to do with whether the cell cycle check-point controls are turned off
during the early cleavage stages of development.
Turning off these controls for a while allows the cell cycle to go much faster,
so the embryo can hurry through the cleavage stages of development.
(with another division every hour, or half hour, or 15 minutes.)
Cell cycle controls are NOT turned off in embryos of humans and other mammals,
So you get 1 cell, 2 cells, 3 cells, 5 cells etc. rather irregularly,
with long times between these divisions, like 16-20 hours.
(which is also a typical cell cycle time for cells in the body, and in tissue culture.)
Once a frog embryo (etc.) has hundreds of cells, the cell cycle controls are Reactivated and the time between cell divisions becomes much longer, and also divisions are no longer synchronous.
More questions for class discussion:
g) Why isn't there as much hurry for mammal embryos, would you guess?
h) Suppose an experimenter removed about half or three quarters of the cytoplasm of a frog egg at the one-cell stage, and then discovered that the division rate slowed down sooner, by one or two cell cycles? Would that imply anything about the process or criteria embryos use to decide when to slow down?
i) What if the same premature slowing of divisions also was the result when researchers injected extra DNA (which they got from bacteriophages! In other words, whatever DNA they just happened to have in their lab + NOT frog DNA)
Gastrulation subdivides each embryo into what have been named the three primary germ layers:
-
Ectoderm
Mesoderm
Endoderm
-
Neural tube ectoderm
Neural crest ectoderm
Somatic ectoderm
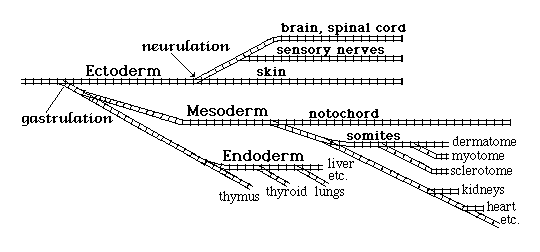
Difficult Question (to which nobody knows the answer):
j) Why use physical rearrangements to decide so many of these branch points, that decide which cell type each embryonic cell will differentiate into?
++++++++++++++++++++++++++++++++++++++++++++++++++++++++++
Some examples of organisms with very different versions of embryology:
Dictyostelium discoideum: a cellular slime mold.
(Considered by many to be a fungus; but we will make it an honorary animal!)
Note that this is just one of many species of slime molds, and there are other genera; but D. discoideum is one of the most intensively-studied of all organisms, because its cells do some important things that human cells also do.
AND this species was discovered by Kenneth Raper, soon after he graduated from UNC back around the 1930s.
-
Dictyostelium aggregation
The surprising life cycle of cellular slime molds:
a) Live as small amoebae, eat bacteria.b) These amoebae divide mitotically.
c) When they run out of food, the amoebae...
-
1) secrete cyclic AMP
2) become chemotactically attracted to cyclic AMP
(so that amoebae attract each other)
3) develop a selective cell-cell adhesion protein
(so that amoebae stick to each other)
(with as many as hundreds of thousands of cells,
or as few as hundreds of cells, or even fewer)
[not the same as the kind of shell-less snail called slugs]
e) These "slugs" crawl around for a few hours
f) Amoebae then differentiate into...
-
* Stalk cells
* Spore cells
* "base cells"
This fruiting body serves as the functional equivalent of a tiny mushroom, holding up the spores so that they can blow away in the air, and land somewhere else.
h) Spores land somewhere, and amoebae hatch out.
Dictyostelium is one of the most widely studied "model" research organisms, with hundreds of research labs around the world concentrating on this one species.
Advantages include:
-
Differentiating into only 4 cell types
Many mutant strains
The form of "amoeboid locomotion" most like our cells'
When you cut the slugs in 2, or 100, they" regulate"
