Lecture notes for Wednesday, Feb. 25, 2009
1) Review of key facts about Teleost fish embryology.
a) Teleost Gastrulation includes one of the most dramatic examples of "Epiboly".i* Epiboly is any spreading of an epithelial cell sheet.
ii* In Teleost development, the whole mass of cells spreads outward and downward away from the animal pole, toward the vegetal pole.
(not just epithelia, but also mesenchyme)
iii* The body axis of the fish is created by active cell rearrangements during this spreading process, with the head forming near what had been the animal pole, and more posterior parts of the body forming one-by-one from head to tail.
iv* External pressure on the top of the "blastodisc" the original sheet of cells will often cause formation of two or three heads, with their body axes converging, so that conjoined twins are formed. The same result can be produced by grafting cells from the developing head area to other locations.
&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&
v*) Teleost embryos develop and depend on two special "Extraembryonic Membranes" = tissues or organs that do not become any part of the eventual body of the animal, exist only during development, and serve either to protect, or provide nutrition, store waste materials, or serve other functions
vi*) The "Enveloping Layer" and the "Yolk Syncytial Layer" are the two extraembryonic membranes of teloeosts, and only they have them.
The enveloping layer is a (very!) thin epithelial cell sheet, that extends over all the rest of the embryonic cells, like a giant protective dome.
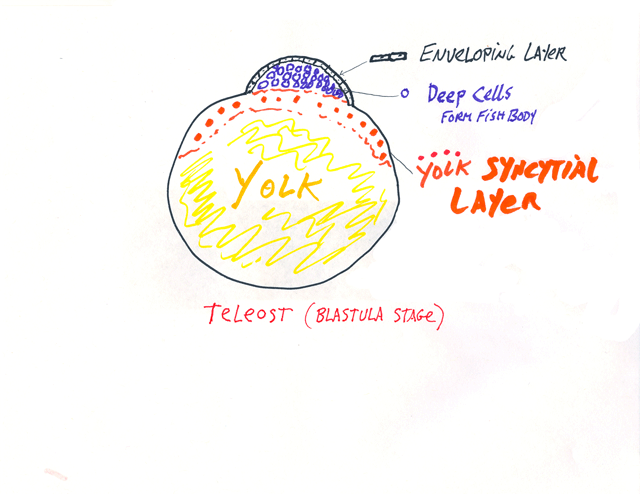
The yolk syncytial layer is below the rest of the embryonic cells,
and consists of many large nuclei, which have failed to separate
into individual cells during cleavage, plus their surrounding cytoplasm.
P.S. For many decades, the yolk syncytial layer was instead named the periblast, because it looked as if it were "around" the blastodisc, but actually extends underneath, and was renamed about 25 years ago. I am afraid that sometimes I forget and use the word periblast. I was against changing the name. But you will not be asked to learn the word periblast, or graded on knowing it. However, knowing this word would help you understand some older (but still important) research papers in journals.
&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&
Amphibian embryos do not have extraembryonic membranes, with a few special exceptions that you don't need to know about. Some other kinds of fish have different extraembryonic membranes than teleosts. Live-bearing teleost fish develop the equivalent of placentas. The details of how they do it are beyond the scope of this course.
Developing embryos of reptiles, birds and mammals have four extraembryonic membranes, and these are parts of the placenta of developing mammals.
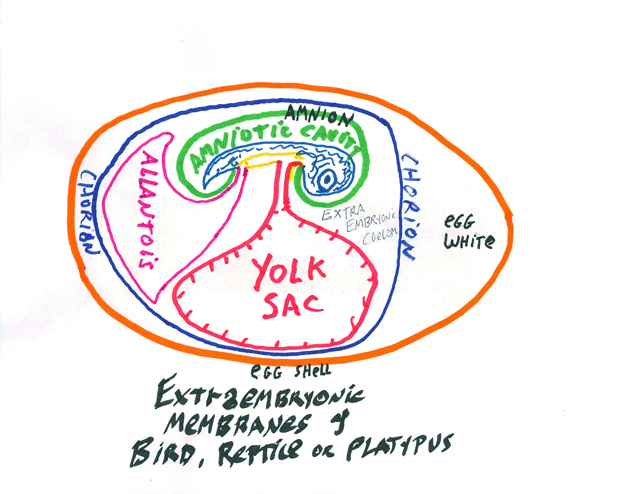
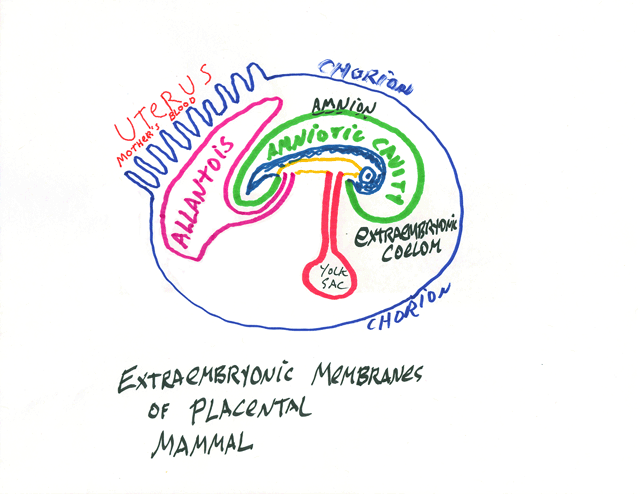
* The Chorion: part ectoderm and part lateral plate mesoderm;
extends around the whole of the embryo, protects it and keeps it wet.
** The Amnion: part ectoderm and part lateral plate mesoderm: Surrounds the developing body, and is colloquially called "The Bag Of Waters"
*** The Yolk Sac: part endoderm and part lateral plate mesoderm: Provides nutrition, and surrounds the yolk, except in mammals. It becomes a side branch of the small intestine.
**** The Allantois: part endoderm and part lateral plate mesoderm; Stores urine; its blood vessels are the main vessels of the placenta, and it develops as a side branch of the bladder.
The Umbilical Cord connects the body to the extrambryonic membranes. (all of which are really organs, please notice, not just membranes.
Turtles, snakes etc. also have belly-buttons, incidentally, where their bodies were connected to their extra-embryonic membranes!
Many snakes are live-bearing: that means that the embryos develop inside the body of the mother snake. Copperheads are one of many examples.
No one knows much about the evolutionary steps leading to the chorion, amnion,etc. In particular, were the intermediate species egg-layers or live-bearers? Either is conceivable There could have been intermediate versions of external eggs in which out-folded epithelial layers gradually surrounded the developing body, more and more completely (culminating in the evolution of the amnion and chorion). Alternatively, these structures could have gradually evolved in live-bearing intermediates between amphibians and reptiles. I don't know what kind of evidence we should look for, either in fossils or in genomes, that could prove or disprove either possibility. A closely related question is whether egg-white and egg shells evolved before or after the chorion, the amnions, and the allantois. I presume that the yolk sac evolved before all of them.
But I would value your opinion on these issues, and about what kind of evidence should be looked for.
Mammalian Embryology Feb 25/27, 2009
From gastrulation onward, the embryology of humans and other mammals becomes nearly the same as in birds.
Before that, most mammal embryos are very different from birds.
Human, mouse (and cow etc.) oocytes are the size of sea urchin eggs.
About a tenth of a millimeter in diameter (100 micrometers)
[nevertheless, that's ~ a thousand times the volume of an
average human cell in the body]
Yolk (stored food material) is evenly distributed through this big cell, which is released from the ovary by rupture of a fluid-filled vesicle (tearing open a new hole in the ovary tissue each time an oocyte is released).
Cycles of ovulation (egg cell maturation, release & first meiotic division)
are controlled by counterbalance of two sets of hormones.
One set, which are large proteins, are secreted by the pituitary,
and stimulate sperm production in males and egg cells in females.
(These are named "Follicle Stimulating Hormone" and "Luteinizing Hormone" = "FSH" and "LH" )
The so-called "fertility drugs" that you hear about in the news are
either analogs of FSH and LH, or maybe these hormones themselves.
By injecting these hormones human ovaries
can be stimulated to produce (ovulate) ~10 oocytes, instead of one.
A sperm is then injected into each of these many oocytes, ("In vitro fertilization") which are then allowed to go through 2 or 3 cleavage cycles outside the body, watched with light microscopes, and the least abnormal 2 or 3 embryos are then implanted into the mother's uterus. The other 7 or 8 developing embryos are then kept (forever) frozen at liquid nitrogen temperatures. (Special freezers are made for this purpose)
Embryos and tissue culture cells are routinely kept frozen this way, and later thawed out and live normally. For example, I have done this hundreds of times with mammal tissue culture cells, using only dry-ice temperatures. There are tricks, but it isn't hard to do.
Steroid sex hormones have the opposite effect (inhibit ovulation), because they inhibit FSH and LH secretion in a negative feedback cycle.
Birth control pills work by this negative feedback, and contain steroids.
The kinds of steroids that are misused by baseball players, etc. also inhibit sperm production in males. Can you figure out how?
______________________________________________________
Sperm travel up the oviducts ("Fallopian Tubules") by a mixture of swimming by means of flagella, and peristalsis by the oviduct.
They meet and fertilize the oocytes at the top of the oviduct.
Cleavage divisions are not synchronous in mammals, occur every 15-20 hours (in contrast to every hour, or 30 min.) and are not synchronous, and have no regular geometric pattern.
Early mammal embryos have what amounts to a shell (rather thick & strong). This is called the zona pellucida in the special case of humans, but for other species it is called the vitelline membrane.
(For historical reasons, especially that different scientists worked on human embryos than worked even on other mammals' embryos, different names are sometimes given to the same structures.)
The textbooks claims (and if you go to med school, you will be required to learn and parrot back on tests) than mammal embryos have "rotational" cleavage, which means that the first four cells get arranged at the corners of a tetrahedron. To the extent that this is true, it is a passive result of the cells being tightly enclosed in their vitelline membrane. It isn't at all comparable to the special cleavage patterns (radial, spiral) that are caused by how mitotic spindles and contractile rings are located. It's an example of third-rate, careerist, semi-scientists advancing their names by "discovering" trivia, and inventing names for them.
For nearly a week, a cleaving human embryo drifts down the oviduct towards the uterus (which is the highly-differentiated, lower end of the oviduct, in which right and left tubes are fused to form one cavity).
After several cleavages, the embryonic cells begin to make adhesion molecules & stick to each other. They also undergo dramatic waves of acto-myosin contractions, as several excellent scientists have studied, but about which the textbook doesn't have a clue. This textbook is the best now available, and has many strengths.
I think it is a very valuable lesson for you students to notice how often what the book says is very different from phenomena that you can plainly see in videos that I show in class. It is a heck of a job for authors to pull together so much information, and inevitable for some to be missed; but I believe Gilbert has a systematic bias against mechanical phenomena and almost no understanding of math or physics.
This stage of cell-cell adhesion and contraction waves
is called "compaction". Sometimes, the cells will compact
together into two spheres, instead of one.
This is one of the most common causes of identical twinning.
Whenever identical twins each have their own set of extraembryonic
membranes, then it can be deduced this is what happened.
After compaction, mammal embryos secrete enzymes that digest the vitelline membrane = zona pellucida, and hatch from it. The embryo then sticks to any nearby tissue, and invades and digests its way down into this tissue, which it stimulates to grow and help form a placenta. This active invasion is called implantation.
Implantation normally occurs in the uterus, which is specialized for it. But compacted and hatched mammal embryos can implant into the wall of the fallopian tubule ("a tubal pregnancy"). Whatever tissue they happen to be next to at the time they hatch, they will implant into it and try to form a placenta.
As the embryo grows inside a fallopian tube, it eventually bursts the tube and dies, after producing enormous pain. Because there is no way to save such an embryo, I do not see how it can be immoral for a surgeon to remove it. Those who get their embryological and moral advice from extremist politicians often object to such things. It is a cheap-shot way for ignorant people to pretend to be more moral.
I should mention that the chance of having a tubal pregnancy is greatly increased by previous scarring of the fallopian tubules by certain sexually-transmitted diseases, especially Chlamydia.
A steroid named progesterol is needed for embryos to implant; a synthetic structural analog of progesterol is used to inhibit implantation and also used to treat certain kinds of cancer.
Soon after compaction, the cells of mammal embryos secrete
water into the spaces between them, so that they inflate like a water balloon.
This stage is called the blastocyst.
This is the closest they come to being a blastula, but is different
because 95-99% of the cells will become parts of the extraembryonic
membranes (collectively, the placenta).
A few interior cells form an inner cell mass across one end of the fluid-filled space inside the blastocyst. The body develops only from cells of the inner cell mass, and even many of these become parts of the placenta.
About half of human identical twins result from formation of two inner cell masses inside the same blastocyst. This mode of twin formation is deduced when twins share the same chorion, but each has its own separate amniotic cavity.
The third, and rarest, cause of human identical twins
is formation of two primitive streaks on the same inner cell mass.
This can be deduced after the fact by the twins being inside
the same amniotic cavity (bag of waters).
Only for this third form of twinning is there danger (possibility) that some parts of the twin's anatomy will be part of both their bodies (conjoined twins = "Siamese twins").
Only a small minority of twins formed this way are conjoined; but none of the identical twins formed in the other two ways are ever conjoined.
Although it is not wrong to say that their bodies are fused, you should not think that separate bodies formed, and then somehow merged. What really happens is that the same mass of cells gets subdivided into the organs of two bodies (as if you were molding two pots from the same piece of clay, would be a possible analogy. I would appreciate being e-mailed any better analogy you can think of.)
For those of you who now love the subject of symmetry, notice that conjoined twins are (nearly?) always mirror images of each other (at least, of those that survive).
This must be telling us some important facts about the normal control mechanisms of body formation. (but nobody knows, or notices).
These are some of the reasons why it is misguided to argue that there is some certain time when an embryo becomes alive. That is about on the level of old Frankenstein movies. Are we going to claim embryos are two people until the latest stage when twinning could occur? And then what? A serious question is: when does a developing embryo develop far enough to have a functioning nervous system, and thus be a person. Sperm are alive; oocytes are alive; they remain alive before and after the second meiotic division, before and after cleavages, compaction, implantation, inner cell mass formation, primitive streak formation, before during and after gastrulation and neurulation.
If somebody knows enough to have an opinion about why conjoined twins are mirror images of each other, then that person gets to have opinions about life beginning. Otherwise, they should try to learn, not be dogmatic, and not be part of hate groups.
Incidentally, even when not conjoined, twins formed from the same inner cell mass are (usually? always?) mirror images of each other. The twin on the right has a heart & intestines with reversed symmetry. The twin on the left is dominant.
I don't see how to explain this by beating of flagella at Hensen's node.
______________________________________________________
Please remember that
*we mammals evolved from egg-laying reptiles,
** three species of mammals remain that still lay eggs,
which are very much like reptile and bird eggs.
(One species of Platypus and two species of Echidnas, in Australia and New Guinea)
Because even Echidnas are so rare and endangered, there has not been much study of their embryology.
Marsupials, like Opossums and Kangaroos have oocytes that are much smaller & less yolky than birds and platypuses, but much larger and more yolky than regular mammals, like us. I think they are an evolutionary side branch & would like to learn more about their embryology, about which there have been many studies. But we have to skip them in this course.
Gastrulation, neurulation and later development are almost the same in regular ("eutherian") mammals as in birds.