February 18th lecture
Embryology Biology 441 Spring 2008 Albert Harris
1) A few more points on Teleost fish embryology.
a) Teleost Gastrulation includes one of the most dramatic examples of "Epiboly".i* Epiboly is any spreading of an epithelial cell sheet.
ii* In Teleost development, the whole mass of cells spreads outward and downward away from the animal pole, toward the vegetal pole.
iii* The body axis of the fish is created by active cell rearrangements during this spreading process, with the head forming near what had been the animal pole, and more posterior parts of the body forming one-by-one from head to tail.
iv* External pressure on the top of the "blastodisc" the original sheet of cells will often cause formation of two or three heads, with their body axes converging, so that conjoined twins are formed. The same result can be produced by grafting cells from the developing head area to other locations.
&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&
v*) Teleost embryos develop and depend on two special "Extraembryonic Membranes" = tissues or organs that do not become any part of the eventual body of the animal, exist only during development, and serve either to protect, or provide nutrition, store waste materials, or serve other functions
vi*) The "Enveloping Layer" and the "Yolk Syncytial Layer" are the two extraembryonic membranes of teloeosts, and only they have them.
The enveloping layer is a (very!) thin epithelial cell sheet, that extends over all the rest of the embryonic cells, like a giant protective dome.
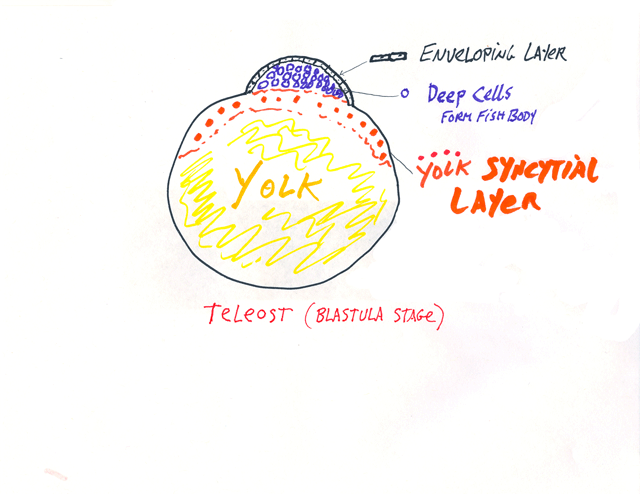
The yolk syncytial layer is below the rest of the embryonic cells,
and consists of many large nuclei, which have failed to separate
into individual cells during cleavage, plus their surrounding cytoplasm.
P.S. For many decades, the yolk syncytial layer was instead named the periblast, because it looked as if it were "around" the blastodisc, But actually extends underneath, and was renamed about 25 years ago.
&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&&
Amphibian embryos do not have extraembryonic membranes. And certain fish have different ones than teleosts.
Developing embryos of reptiles, birds and mammals have four extraembryonic membranes, and these are parts of the placenta of developing mammals.
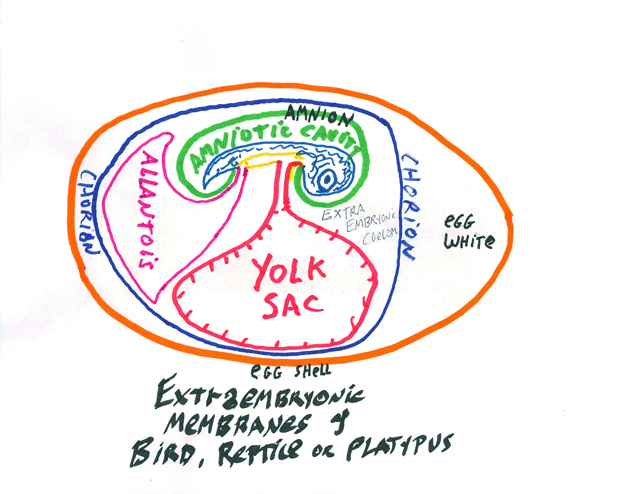
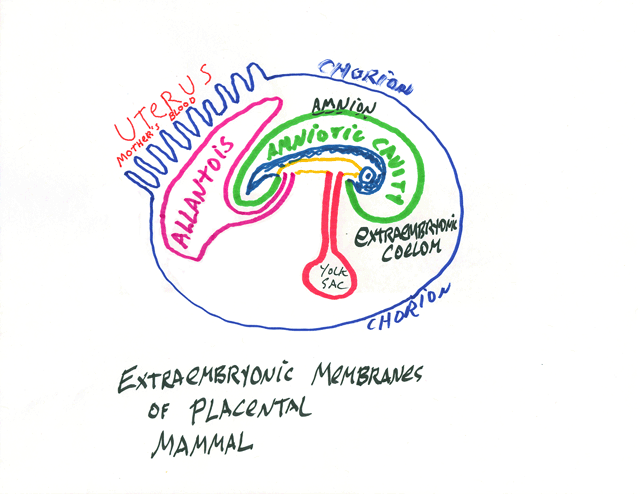
* The Chorion: part ectoderm and part lateral plate mesoderm;
extends around the whole of the embryo, protects it and keeps it wet.
** The Amnion: part ectoderm and part lateral plate mesoderm: Surrounds the developing body, and is colloquially called "The Bag Of Waters"
*** The Yolk Sac: part endoderm and part lateral plate mesoderm: Provides nutrition, and surrounds the yolk, except in mammals. It becomes a side branch of the small intestine.
**** The Allantois: part endoderm and part lateral plate mesoderm; Stores urine; its blood vessels are the main vessels of the placenta, and it develops as a side branch of the bladder.
The Umbilical Cord connects the body to the extrambryonic membranes. (all of which are really organs, please notice, not just membranes.
Turtles, snakes etc. also have belly-buttons, incidentally, where their bodies were connected to their extra-embryonic membranes!
In this course, you learn something new every day.
Back to Teleost fish embryonic development (sort of)
*****************************************************************
H.V. Wilson wrote the first complete description of how teleosts develop, specifically using eggs of the Black Sea Bass, at Woods Hole.
That research was what earned his appointment as Professor at UNC, where he began the course you are taking, in the late 1890s.
His most important discovery was sponges and corals can re-form functioning individuals even after all their cells have been dissociated and randomly mixed back together. ("cell sorting")

John Phillip Trinkaus, also working at Woods Hole, became the leading Teleost Embryologist of the 20th century, working on killifish.
Trinkaus was one of the first to trace cell fates with radioactive isotopes, first during his own PhD research on neural crest in chickens. Later he used tritiated thymidine to prove that embryonic cells of mice and birds can also sort out after being dissociated into random mixtures of individual cells. They don't reform animals, but sort out by differentiated cell type, for example liver cells sort out from heart cells to form internal blobs of heart cells surrounded by continuous layers of liver cells.
Chick, mouse and amphibian embryonic cell don't reform animals, but sort out by differentiated cell type, for example liver cells sort out from heart cells to form internal blobs of heart cells surrounded by continuous layers of liver cells.
Other major researchers on this subject were Johannes Holtfreter, Aaron Moscona and Malcolm Steinberg.
Using Amphibian embryos, Holtfreter discovered that their cells will sort out from random mixtures according to germ layer, and subdivisions of germ layers.
For example, he found that neural ectoderm will sort out from somatic ectoderm, and that mesoderm will sort out from ectoderm and endoderm. Mesoderm acquires the ability to sort out at the time of gastrulation.
Holtfreter interpreted these results by a theory of changing "tissue affinity", which led to the discovery of cell-cell adhesion proteins, like the cadherins. Research by Rachel Fink at Duke confirmed that rearrangements of cells inside embryos can be caused by changes in which cadherins are on the ouside surfaces of particular cells.
Now, back to Teleost Embryos
The neural tube of teleosts forms as a solid rod of cells that (by some unknown mechanism) hollows itself out.
(Instead of a sheet of cells rolling up to form a tube, like in bird, mammal, amphibian gastrulation!?)
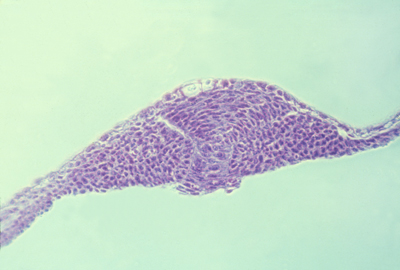
This must mean something: What does it mean?
Does it mean that fish use an entirely different mechanism to form their nervous system?
Or does it mean that all vertebrates use the same mechanism, that causes sheet to fold AND causes solid rods to "cavitate"?
If you dissociate neural plate cells (of birds, amphibians, etc.) they first reaggregate to form solid masses that then undergo cavitation, to form neural tubes.
This is only one of many examples in which embryos seem to use qualitatively different mechanisms to form particular organs.
Some theories about how embryonic cells produce the same end results (same geometry and patterns)
1) Steinberg's Thermodynamic theory of cell sorting. (Cells maximize their cell-cell adhesions)
2) Ingber's tensegrity theory. Cells arrange so that there are exact counter-balances of pulling and pushing forces.
3) The Theory of Anatomical Homeostasis. Cell traction and other forces adjust relative strengths, as functions of shape and arrangement, so that different geometrical arrangements become stable.
As body temperature becomes 98 degrees, and is kept at that temperature, because more sweating occurs at higher temperatures, and more shivering occurs at lower temperatures.

I realize this doesn't directly explain fish neurulation, etc. The more general questions are:
A) How do changes in cell properties cause them to arrange into normal anatomical patterns?
B) Why can cells reach the same eventual geometrical arrangements by multiple alternative sequences of intermediates?
C) Can the mechanisms that cause cell arrangements be controlled for medical purposes?
To guide wound repair, for plastic and orthopedic surgery, or even to regenerate organs...