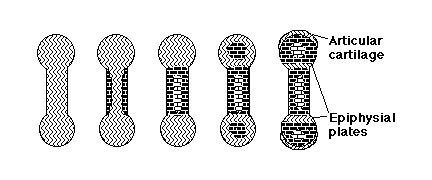Embryology Biology 441 Spring 2010 Albert Harris
Lecture Notes for March 19
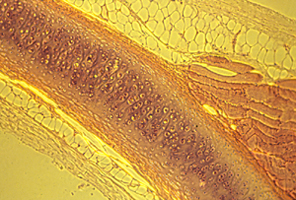
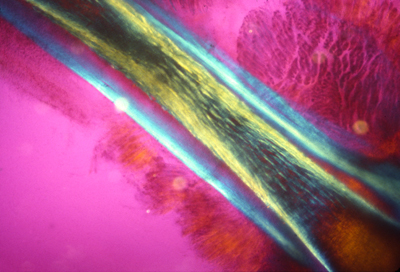
Embryonic artery, wrapped in collagen fibers (yellow)
First sub-topic, blood vessels: which are formed from mesoderm.
Capillaries; Veins; Arteries; + also Lymphatic ducts.
In all of these, the innermost lining is a sheet of a special kind of epithelial cell, called "endothelial cells" "endothelium"
(Don't be fooled by the name, these are NOT formed by endoderm!)
Capillaries are narrow hollow tubes of endothelial cells, with little or no other reinforcement. Despite their very thin walls, the pressure in capillaries is more than in veins, and in many capillaries the pressure is almost as strong as in arteries!!?
(90+% of biologists don't really understand how this is possible!)
But earlier in the course you learned about P = C*T + c*t
Capillaries have very high Curvature, around their circumference.
So Pressure can be very large, & Tension is not too strong.
(The same principle is used in backhoes and other heavy equipment
Sometime, notice the thin pipes that carry their high pressure fluid)
Veins are much wider tubes, in which the inner-most lining is a tube of endothelial cells, but they are surrounded by many layers of fibroblasts (mesenchymal cells), and wrapped by Type I collagen fibers (which are mostly secreted by the fibroblasts).
Arteries: innermost layer of endothelial cells
Wrapped around that is a thick, strong layer made mostly of smooth muscle cells, with their long axes, and directions of contraction, oriented in the circumferential direction.
Also fibroblasts and collagen fibers are wrapped around circumferentially; plus another fibrous protein "Elastin"
Elastin is rubber-like, 'stretchy'; In contrast to collagen which forms strong nylon-like fibers and sheets.
Veins also have some elastin.
The aorta differs from other arteries in that its walls have many fewer smooth muscle cells, and many more fibroblasts, and LOTs more elastin, along with the collagen fibers.
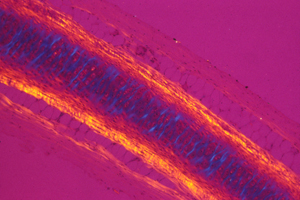
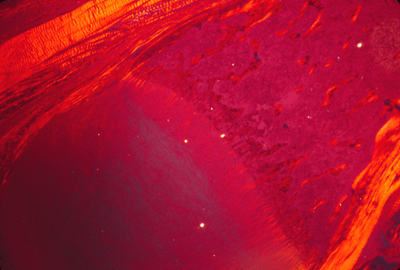
Here is a photograph I took of a cross section through a vein and an artery. (a normal artery)
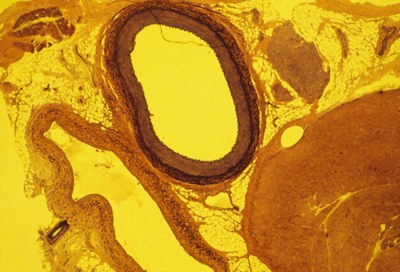
The artery is in the upper-middle.
The vein is below it and somewhat to the left.
The large solid red blob in the lower right is a skeletal muscle.
The smaller red blob to the right of the artery is a bunch of nerves
One of the biggest causes of human death and disability is the disease "atherosclerosis". It is true that if the concentration of cholesterol in your blood is high, then you are more likely to have your arteries blocked by atherosclerotic "plaque".
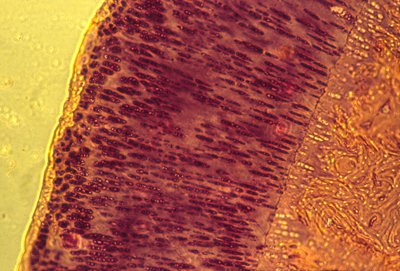
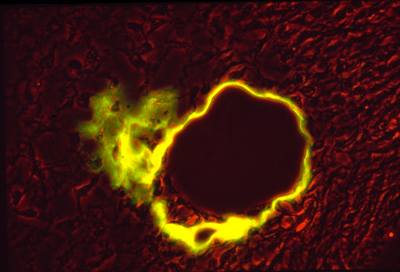
A cross section of a human artery blocked by atherosclerosis.
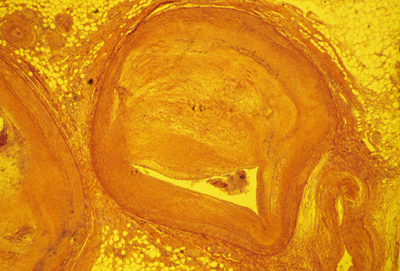
(with another blocked artery to the left, and a vein to the lower right)
NOTICE THAT THE BLOCKAGE CONSISTS MOSTLY OF TOO MANY SMOOTH MUSCLE CELLS,
and that these are located outside the endothelium tube.
It is definitely NOT true that cholesterol sticks to the inside of anyone's arteries. Clotted blood & blood platelets do stick there, and cholesterol accumulates in or among the cells that are blocking the artery.
(some clotting is easily visible in this section)
WHAT IS THE TRUE STORY:
* It IS TRUE that if your body has too much cholesterol, then you will get more and worse arterial blockages of this kind!
* Among all those extra smooth muscle cells, that are bulging into the space where the blood should flow, cholesterol does accumulate. So there is some kind of connection.
But many of you will be going into medicine or medical research, so you should be told the truth, not Aesop's fables, and in this case the truth is that nobody really understands what's happening.
One theory: The blockages are tumors of the smooth muscle cells, analogous to many small cancers, caused by somatic mutations.
Another theory: Turbulence in the blood stimulates blood platelets to secrete PDGF (Platelet-Derived Growth Factor), a protein which is known to stimulate smooth muscle cells to grow and divide and also is a chemotactic attractant for them.
Notice that neither of these theories predicts the (true) fact that these over-growths of smooth muscle cells are larger and worse in animals and people who have more cholesterol in their blood.
If you could invent some possible reasons, and also how to test them, then that would be a HUGE contribution to medical science.
Research science is full of riddles and paradoxes of this kind.
Usually you can't just observe complex phenomena.
Almost always, you have to INVENT THEM, and then look for predictions (surprising predictions will convince people more)
Inventing theories tells you what details to be curious about!
The following are entirely imaginary; there is no evidence for them. (yet)
-
1) When the smooth muscle cells move and divide in response to the PDGF, then
cholesterol precipitates between them, and either makes them grow or move more,
or maybe it somehow locks them in position.
2) Cholesterol deposits make smooth muscle cells more sensitive to PDGF.
3) Cholesterol stimulates platelets to secrete more PDGF, and it's just a coincidence that cholesterol also gets deposited in among the extra smooth muscle cells.
4) Cholesterol deposits prevents apoptosis of extra smooth muscle cells, so that excess accumulations of smooth muscle are not eliminated..
5) Cholesterol somehow causes or allows smooth muscle cells to pull more strongly on collagen fibers, or elastin fibers, or to contract more strongly.
6) Cholesterol deposits between smooth muscle somehow stimulates them to divide.
7)
8)
9)
10) please invent more theories!!!
Most of these are in the yolk sac, in birds and mammals.
Maybe that's why mammal embryos continue to have yolk sacs?
Red blood cells and white blood cells differentiate inside;
Endothelial cells crawl and connect other to form hollow tubes: the first capillaries. Only much later does bone marrow become the place where blood formation occurs
Early embryos don't have bones; and when the skeleton does form it is nearly all made out of cartilage, which is solid, without a marrow, and isn't penetrated by blood vessels ("vascularized")
The yolk sac is one of several different organs that are used as locations for the hemopoietic stem cells which form red blood cells (and also white blood cells). These stem cells move to the liver, and later to the bone marrow.
Embryonic and fetal hemoglobins are different from (and coded for by different genes than) the hemoglobin we have in the red blood cells we make after birth. Fetal hemoglobin binds oxygen slightly more strongly than adult hemoglobin, which increases the amount of oxygen that is transferred across the placenta from the blood of a pregnant woman to the blood of the fetus developing inside her. Two other interesting aspects are:
#1) The genes for embryonic and fetal hemoglobin proteins are directly adjacent to the genes for adult hemoglobins, with the early blood cells activating one gene, and then later blood cells activating the gene next to that one. The mechanism isn't known, nor whether there is any similarity to the mechanisms that cause adjacent Hox genes to be transcribed in adjacent tissues.
#2) A few people continue to make fetal hemoglobin all their life, instead of switching to the adult hemoglobin genes, and the symptoms of this are almost unnoticeable! So far as I know, it hasn't yet been discovered what mechanism causes this; but it could become a new and better treatment for sickle-cell anemia (and other genetic defects of adult hemoglobins), if you could discover a drug or other treatment that switches their hemopoietic stem cells back to transcribing the genes for fetal hemoglobin.
The Skeleton: formed by mesodermal cells,
except the face, where skeleton is neural crest cells
>
Vertebrate skeletons are made partly of bone and partly cartilage.
Bone compared with Cartilage
Bone is stronger and more rigid,
but can grow only by surface deposition.
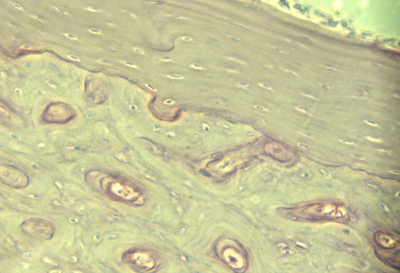
cross section of bone
Cartilage is lighter, flexible, weaker,
but can grow by internal swelling (& also by surface addition).
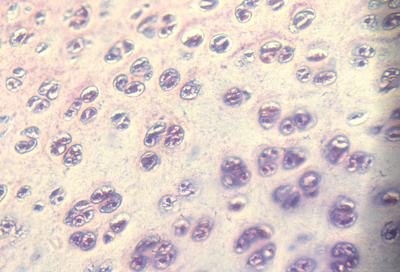
cross section of a typical cartilage
Bone is a tightly woven composite of 1/3 type I collagen fibers,
embedded among crystals of calcium phosphate (2/3rds by weight)
(& also fluoride and hydroxide) (dentine is almost the same)
If horses are "sent to the glue factory", it's for bones not hoofs.
Bone is made (but not really secreted!) by osteocytes. (= osteoblasts)
Deposition of bone is called "ossification".
Osteocytes do secrete collagen, but I think it is still an unsolved problem exactly what mechanism they use to create the calcium phosphate crystals!
Another special differentiated cell type (osteoclasts) constantly dissolves and destroys bones. Bone is a very dynamic tissue, being constantly
destroyed and remade. When osteocytes don't keep up with the osteoclasts
the result is osteoporosis. There are many unsolved problems here.
Is it over-activity of osteoclasts? Or underactivity of osteocytes?
How can you prove which is the reason? How to cure it?
Many drugs are designed to strengthen the calcium salts, but these drugs can have very painful and harmful side effects (do a web search and read the horrifying reports from patients). A possible explanation is that these "bone strengthening" drugs induce programmed cell death (apoptosis) in the osteoclasts (bone digesting cells) and also some or many other cell types such as in the digestive tract.
Cartilage is a mixture of type II collagen and chains of sugar molecules that have sulfate groups covalently bound to them. The sulfates ionize,
and their negative charges keep a cloud of cations (like Na+) nearby.
This excess concentration of cations causes an osmosis-like inflow
of water ("electro-osmosis"), and this swelling pressure of water is what makes cartilages stiff and makes them tend to expand.
(Very few biologists understand the physics of cartilage swelling!)
Cartilages grow partly by increased synthesis of sulfated sugars, and partly by selective weakening of both the internal and surface collagen.
As an analogy, imagine controlling the shape of a balloon partly by forming more gas inside, and partly by weakening the rubber.
There is a big need for better research in this whole area.